Sleep stages are the four distinct neurophysiological states the brain cycles through every night, identifiable from the EEG, the EOG (eye movements), and the EMG (muscle tone), and they are the thing the entire sleep-and-cognition literature is actually talking about when it says things like "you need more deep sleep" or "fragmented REM tanks emotion." Total sleep duration is the headline metric most people track, but duration is a proxy for what the stages collectively deliver: about a quarter of the night in slow-wave sleep doing procedural and declarative memory consolidation plus glymphatic clearance, about another quarter in REM doing emotional regulation and episodic integration, with the rest in lighter NREM stages doing sleep-spindle-mediated consolidation and homeostatic transition work. Knowing the stages turns sleep from "how many hours" into "what work my brain actually got done last night."
This piece is the cognitive-science version of sleep architecture: the four sleep stages (N1, N2, N3, REM), the EEG signatures that distinguish them, the ~90-minute ultradian cycle that runs four to six times per night, the within-night progression that front-loads slow-wave sleep and back-loads REM, what each stage actually does at the neural and behavioural level, the difference between sleep latency, sleep efficiency, and sleep fragmentation, and why a 7-hour night with fragmented architecture often produces worse next-day focus than a 6-hour night with intact cycles. Tomatoes is a focus tool built around the daytime side of this: the working block where the cognitive operations sleep was supposed to consolidate get used. The app is free for 3 days, then $4.99/week, $29.99/year, or $39 lifetime.
What Sleep Architecture Actually Is
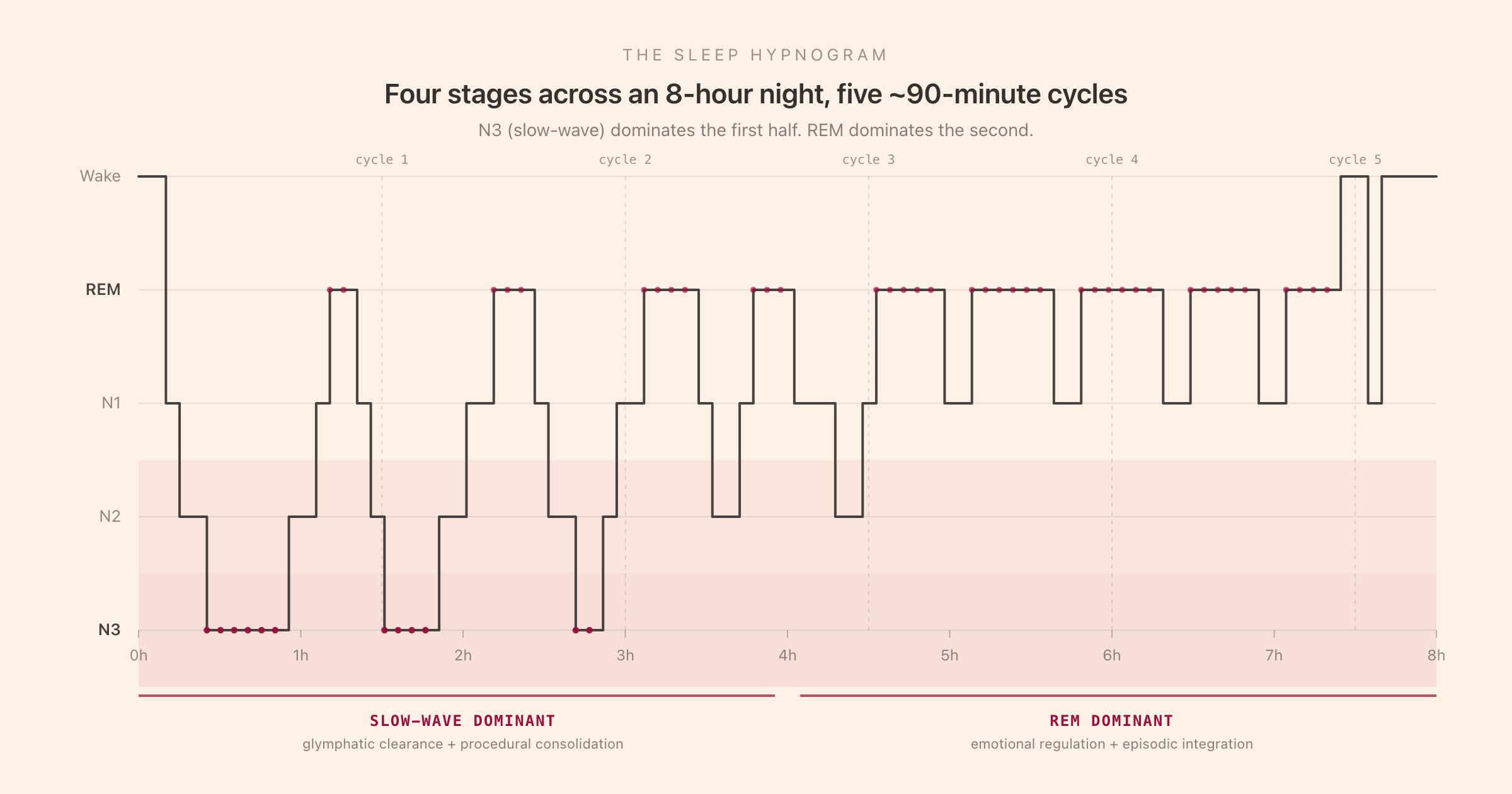
Sleep architecture is the term for the temporal organisation of the night: which stages occur, in what order, for how long, and how often. The architecture is visible on the hypnogram, the stepped-line plot above that shows stage on the Y axis and time on the X axis. A hypnogram is what comes out of an overnight polysomnography study, and it is the canonical way the sleep-medicine literature represents a single night.
The stages themselves are defined by the American Academy of Sleep Medicine (AASM) scoring rules, which classify each 30-second epoch of the night into one of five states from the combination of EEG, EOG, and EMG channels. The five-state classification is Wake plus four sleep stages: N1, N2, N3 (the three NREM stages), and REM (rapid eye movement). The older Rechtschaffen and Kales (1968) system split slow-wave sleep into a separate N3 and N4; the AASM merged them in 2007. Most contemporary literature uses the four-stage AASM system.
The stages are real neurophysiological states, not arbitrary cut-points. The brain in N3 is doing fundamentally different work to the brain in REM, with different dominant brainwave frequencies, different neuromodulatory states (cholinergic activity peaks in REM, drops in NREM), different muscle tone (active in NREM, paralysed in REM via brainstem atonia), and different metabolic and gene-expression profiles. The architecture is the schedule by which the brain spends time in each state.
The Four Sleep Stages
N1: the light-onset stage
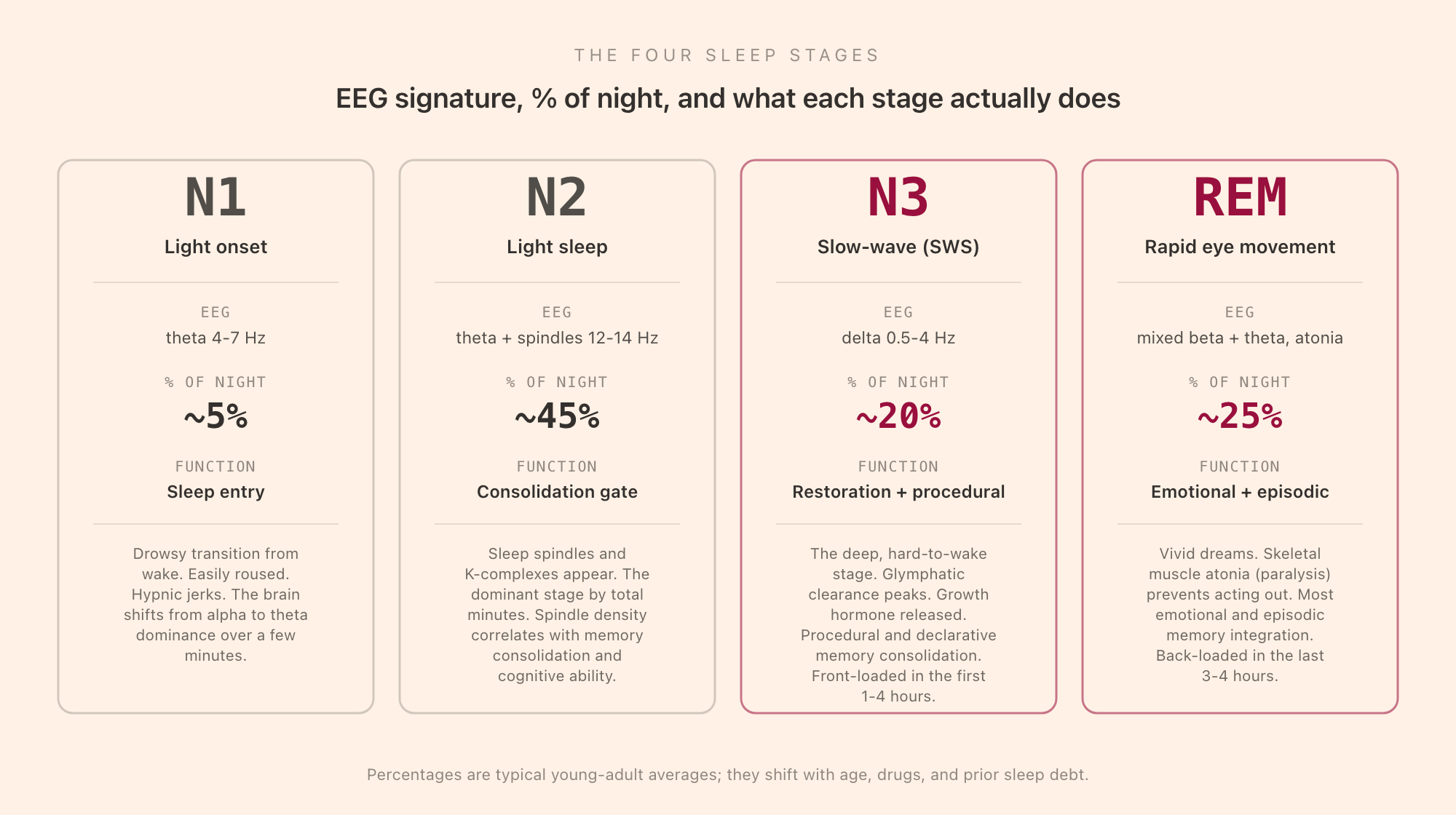
N1 is the transition from wake into sleep. The EEG shifts from the alpha rhythm (8-12 Hz, eyes-closed wake) to a low-amplitude mixed-frequency pattern dominated by theta (4-7 Hz). Slow rolling eye movements appear on the EOG. Muscle tone is still present but reduced. N1 is the stage of hypnic jerks (the involuntary muscle twitches that sometimes wake you as you fall asleep) and of dream-like microimagery that does not yet have the full narrative structure of REM dreaming.
N1 makes up about 5% of a typical young-adult night, and almost all of it occurs at the very start (the few minutes between lights-out and the first sustained sleep) and at brief re-entries after micro-arousals during the night. It is the easiest stage to be woken from; many people awakened during N1 will insist they were not actually asleep, and they are partially right. N1 is the gateway, not the destination.
N2: the dominant stage and the consolidation gate
N2 is the largest stage by total minutes, occupying around 45% of a typical night. The EEG continues to show theta dominance but now adds two diagnostic graphoelements: sleep spindles (brief 11-15 Hz bursts lasting 0.5 to 2 seconds) and K-complexes (single high-amplitude biphasic waves). Spindles are generated by the thalamic reticular nucleus and are increasingly understood as the active mechanism by which N2 supports memory consolidation. Higher spindle density in N2 correlates with better next-day procedural performance, with measures of general cognitive ability, and with the consolidation of newly-learned material across the night.
N2 is the cruise altitude of the night. The brain is unambiguously asleep (muscle tone is low, arousal threshold is up), but the EEG is still relatively active compared to the deep slow-wave stage that follows. N2 episodes are interspersed throughout the night, between bouts of N3 in the first half and between bouts of REM in the second.
N3: slow-wave sleep, the restorative deep stage
N3 is the stage everyone means when they say "deep sleep." The EEG is dominated by delta waves (0.5-4 Hz, high amplitude), and the AASM scoring rule requires more than 20% of the 30-second epoch to be delta-dominated to count as N3. Slow-wave sleep occupies about 20% of a typical young-adult night, and it is the most temporally clustered stage: most N3 occurs in the first 1-4 hours after sleep onset, with much less in the second half of the night.
N3 is where the literature on the restorative and clearance functions of sleep lives. Glymphatic flow (the brain's waste-clearance system, which uses the cerebrospinal-fluid pathways around the brain's blood vessels) peaks during slow-wave sleep, clearing metabolic byproducts including beta-amyloid that accumulate during waking activity. Growth hormone secretion peaks during N3 in the first sleep cycle. Procedural memory consolidation (motor skills, sequence learning) is associated with slow-wave activity, as is the systems consolidation of declarative memories from hippocampus to neocortex.
It is also the hardest stage to be woken from. Someone aroused during deep N3 will be disoriented for several minutes (sleep inertia), which is why an alarm that fires during early-night N3 produces the worst morning grogginess. The reason sleep-stage-targeted alarm apps appeared in the 2010s is that the cost of being woken from N3 is genuinely high; the question of whether consumer apps can detect the stage well enough to time around it is separate.
REM: rapid eye movement, the dream stage
REM is the most distinctive sleep stage. The EEG looks paradoxically wake-like (mixed beta and theta, low amplitude), the EOG shows the rapid eye movements the stage is named for, and the EMG drops to near-zero in a state called REM atonia that paralyses most skeletal muscles except the diaphragm and the small eye muscles. The atonia is generated by brainstem nuclei (the subcoeruleus area in the pons) and is the mechanism that prevents people from acting out their dreams; the rare disorder REM Behaviour Disorder (RBD) results from atonia failure.
REM occupies about 20-25% of a typical young-adult night and is back-loaded: the first REM episode is brief (5-10 minutes) and occurs about 90 minutes after sleep onset; later REM episodes lengthen, with the final episode of the night sometimes running 30 minutes or more. This is why people who wake naturally in the morning often wake out of REM with a vivid dream still in mind.
REM is the stage most associated with emotional regulation and episodic memory consolidation. Functional imaging during REM shows high activity in the amygdala and limbic system with reduced activity in the dorsolateral prefrontal cortex, which is part of the mechanism by which REM helps process emotional content from the day. Selective REM deprivation (waking people every time they enter REM, but allowing N1 to N3) produces measurable deficits in mood, in emotional-content memory, and in creative problem-solving. The cholinergic system (acetylcholine) is highly active during REM, which is part of the pharmacological basis for REM dreaming and for the cognitive operations REM performs.
The 90-Minute Ultradian Cycle
The four stages do not occur in random order. They cycle through a ~90-minute ultradian rhythm, with each cycle progressing roughly: brief N1, then a longer N2, then a bout of N3 (especially in the first cycles), then back through N2, then REM. A typical young adult will run four to six such cycles per night, depending on total sleep duration: four cycles fit in roughly 6 hours, five in 7.5 hours, six in 9 hours.
The cycle length is approximate. Individual cycles can run from 70 to 110 minutes, and the composition of each cycle changes through the night. The first cycle is N3-dominant, with a long bout of slow-wave sleep and a brief, sometimes-skipped first REM episode. Middle-of-night cycles are more balanced, with shorter N3 bouts and longer REM episodes. Late-night cycles are REM-dominant, with little to no N3 and extended REM episodes that can dominate the entire 90 minutes.
This is what produces the front-loading of N3 and the back-loading of REM that the hypnogram makes visible. If you cut a night short by waking up 90 minutes early, you are not losing 90 minutes of evenly-distributed sleep; you are losing a disproportionate amount of REM. If you cut a night short at the beginning (going to bed 90 minutes late), you are losing a disproportionate amount of N3. These are different losses with different next-day consequences.
The 90-minute cycle is one of the body's ultradian rhythms, the family of biological rhythms shorter than 24 hours that the brain runs throughout the day and night. The same approximate cadence appears in daytime alertness curves (the BRAC, basic rest-activity cycle) and in the periodic dips in concentration most people experience every hour and a half.
What Each Stage Actually Does
The functions of the stages are unevenly distributed and reasonably well-characterised in the contemporary literature.
Memory consolidation is split across N3 and REM. N3 is associated with consolidation of declarative memory (facts and events) and procedural memory (motor skills). The thalamocortical slow oscillations that define N3 are thought to coordinate hippocampal sharp-wave ripples with cortical replay, transferring newly-encoded memories from short-term storage to long-term distributed cortical representations. REM is associated with consolidation of emotional and procedural memory, with insight and creative recombination, and with the integration of new memories into existing semantic networks.
Glymphatic clearance peaks during N3. Maiken Nedergaard's group showed in 2013 (Xie et al., Science) that the interstitial space in the mouse brain expands during sleep, increasing the rate at which cerebrospinal fluid washes through and clears metabolic waste including beta-amyloid. The clearance is most active during slow-wave sleep. Chronically inadequate N3 is now suspected as one of the risk factors for neurodegenerative disease, including Alzheimer's, though the causal direction is not fully settled.
Emotional regulation is REM-dependent. REM deprivation produces shifts in emotional tone the next day, with stronger reactivity to negative stimuli and weakened decoupling between amygdala and prefrontal regulation. Walker's "Why We Sleep" (2017) summarised much of this literature; later replications have been mixed on specific claims but the broad finding holds.
Growth and immune function peak in early-night N3. Growth hormone is released in pulses during the first slow-wave sleep cycle. Cytokine activity that supports immune function rises during early-night N3 as well. These are reasons why the first 3-4 hours of the night, where N3 clusters, are not interchangeable with the last 3-4 hours.
Synaptic homeostasis runs throughout NREM. The synaptic homeostasis hypothesis (Tononi and Cirelli) proposes that wake produces a net increase in synaptic strength across the day, and that NREM (especially slow-wave) acts to renormalise that strength downward overnight, freeing capacity for new learning the next day. This is part of why sleep deprivation produces difficulty learning: the homeostatic reset is incomplete.
Sleep Latency, Sleep Efficiency, and Sleep Fragmentation
Three sleep-medicine concepts that the consumer literature mixes up regularly.
Sleep latency is the time from lights-out (or from getting into bed with the intent to sleep) to the first epoch of sustained sleep, typically the first epoch of N2. Normal sleep latency is roughly 10-20 minutes. Latency under 5 minutes is a marker of significant sleep debt or of a sleep disorder (the multiple sleep latency test, MSLT, uses very short latencies as a diagnostic for narcolepsy). Latency over 30 minutes consistently is one of the diagnostic criteria for insomnia.
Sleep efficiency is the ratio of total sleep time to time in bed. A person who is in bed for 8 hours and sleeps for 7.5 has 93.75% sleep efficiency. Normal sleep efficiency is around 85-95%; below 85% sustained is a marker for insomnia, sleep-disordered breathing, or fragmentation from external causes. Efficiency is the metric most consumer wearables get closest to estimating accurately, because total sleep time is easier to detect from movement and heart rate than specific stages are.
Sleep fragmentation is the rate of arousals or transitions out of sleep during the night. The arousal index (AI), measured in formal polysomnography, counts brief EEG arousals per hour of sleep. Normal AI is under 10/hour; over 20/hour is significantly fragmented and is a marker of disorders like obstructive sleep apnea or periodic limb movement disorder. Fragmentation matters because each arousal can break a sleep cycle mid-stage, preventing the brain from completing its N3 or REM work even if total sleep time looks normal on a wearable.
The practical implication: a 7-hour night with 30 arousals an hour can produce more next-day cognitive deficit than a 6-hour night with 5 arousals. Total time is a necessary but not sufficient sleep metric; architecture is the underlying variable.
Why Fragmented Sleep Tanks Next-Day Focus
The link between fragmented sleep and next-day focus is mechanistic, not just correlational.
When sleep fragments, the brain spends less time in completed cycles and more time in N1 and N2. Both N3 and REM require sustained, uninterrupted sleep to enter and to remain in. A fragmented night will skew the architecture toward light sleep at the expense of the two consolidation-heavy stages. The next-day costs show up in:
- Working memory (the four-slot cognitive bottleneck that runs every focus task) is degraded after fragmented sleep even when total sleep time is normal.
- Executive function (the prefrontal control system that decides whether to focus) is degraded, particularly on the inhibition and shifting subcomponents.
- Emotional regulation is degraded because REM was reduced; small frustrations during the day produce outsized responses.
- Procedural memory consolidation is incomplete because N3 was reduced; skills practised the previous day do not show the typical overnight improvement.
- Sustained attention is degraded across the day, often with subjective alertness that does not match objective performance (you feel fine but make more errors).
This is the mechanistic basis for brain fog of the sleep-driven subtype. The driver is not duration; it is the architecture failing to deliver enough completed cycles of N3 and REM.
How Caffeine, Alcohol, Age, and Drugs Shift the Hypnogram
The architecture is sensitive to common substances and to age in well-characterised ways.
Caffeine delays sleep onset, reduces total N3, and increases night-time arousals. Caffeine's half-life is 5-6 hours in most adults; a 200 mg coffee at 4pm leaves about 100 mg in the bloodstream at 10pm, still enough to push the first slow-wave bout later and shallower. The effect is dose-dependent and individual (CYP1A2 metabolism varies); slow metabolisers can show effects from caffeine taken before noon.
Alcohol is a famous architecture-disruptor. The first half of the night after alcohol shows MORE N3 (alcohol is a sedative), but the second half shows compensatory REM rebound with vivid dreaming and frequent awakenings, plus rebound autonomic activation. Net effect: total sleep time may be similar but the architecture is markedly worse, with reduced restorative quality.
Age progressively reduces N3. A typical young adult might get 100 minutes of N3 a night; a healthy 65-year-old might get 30 minutes; an 80-year-old often gets close to zero. REM is more preserved across age but also reduces gradually. Total sleep time is reasonably stable across adulthood; the underlying architecture is not.
SSRIs and many other antidepressants suppress REM, particularly in the first weeks of treatment. The effect is part of how some antidepressants work (REM suppression is associated with mood improvement in some models) but it changes the architecture significantly.
Benzodiazepines and Z-drugs (zolpidem, eszopiclone) increase total sleep time, but suppress N3. Net effect: longer time asleep, less restorative quality. The chronic-use consequence is one of the reasons sleep medicine treats these as short-term solutions, not maintenance therapy.
How Sleep Architecture Connects to Daytime Focus
The architecture-to-focus link runs through several of the systems Tomatoes is built to support during the day.
The working block presupposes a well-architected previous night. A focus session draws on working memory, executive function, sustained attention, and emotional regulation. Each is degraded by inadequate N3 or REM. The single biggest performance lever for sustained focus during the day is the quality of the prior night, not the quality of the music or the technique in the moment.
The ultradian rhythm continues during the day. The same ~90-minute cycle that runs four to six times overnight continues to run as the daytime Basic Rest-Activity Cycle (BRAC) Nathaniel Kleitman first described. Daytime alertness peaks for roughly 70-90 minutes, then drops for 15-20 minutes, then peaks again. The pomodoro cadence (25/5 or 50/10) is a coarse approximation to this; the longer 90/20 working-block cadence is a closer fit. Both ride on architecture that the brain established overnight.
Strategic napping uses the same stages. A 20-minute nap is short enough to stay in N1 and N2 (no N3), so you wake without sleep inertia and get a small alertness bump. A 90-minute nap is long enough to complete one full cycle including N3 and REM, so you wake from light sleep at the end of the cycle and get more substantial cognitive restoration. A 45-minute nap is the worst-of-both: it lands in mid-N3, so you wake disoriented from sleep inertia without having completed a cycle. Nap design is sleep-architecture design.
How Tomatoes Fits
Tomatoes does not replace sleep, and it does not claim to influence sleep architecture. What it does is fit the daytime side of the architecture: it provides the audio scaffolding (binaural beats, brown noise, granular textures, the 40 Hz gamma and theta-band options) for sustained working blocks that draw on the working memory, executive function, and sustained attention the previous night's N3 and REM are responsible for consolidating and maintaining.
The connection is real but indirect. You cannot focus-music your way out of a fragmented night; the cognitive substrate is degraded and no audio will repair it. What you can do is build the daytime focus block to match the architecture you actually have. On a good-architecture night, a 90/20 cadence with sustained audio scaffolding gets the most out of a high-functioning prefrontal cortex. On a fragmented night, dropping to a 25/5 cadence with more frequent recovery and lower ambition is the more honest match.
Tomatoes is free for 3 days, then $4.99/week, $29.99/year, or $39 lifetime. It runs locally as a desktop app with a system menu-bar companion, generates the audio in real time, and is designed for working blocks of three to six hours per day, the daytime side of a four-to-six-cycle night.
The Short Version
Sleep architecture is the temporal organisation of the night across four stages (N1, N2, N3, REM), cycling on a ~90-minute ultradian rhythm four to six times a night, with slow-wave sleep front-loaded in the first half and REM back-loaded in the second. Each stage does specific work: N3 handles glymphatic clearance and procedural and declarative consolidation, REM handles emotional regulation and episodic integration, N2 acts as the spindle-mediated consolidation gate that occupies almost half the night, N1 is the brief transition. Sleep latency, sleep efficiency, and sleep fragmentation describe the timing and integrity of the architecture; fragmentation tanks next-day cognition even when total sleep time looks normal. Caffeine, alcohol, age, and many medications shift the architecture in specific ways. The daytime focus block is what gets done with the cognitive substrate the previous night's architecture either built or failed to. Total hours is a coarse proxy; what your brain actually got done last night is the stages.