Norepinephrine is the brainstem neurotransmitter that decides whether attention works. It is released from a single tiny nucleus in the pons (the locus coeruleus, the "blue spot") whose neurons project to almost the entire cortex, and the firing pattern of those neurons sets the gain on every cognitive system downstream. When the locus coeruleus fires in clean, event-locked bursts on a low background, the brain is focused. When it fires at a high sustained baseline with no bursts, attention slips and the same person becomes distractible. The same molecule, the same nucleus, the same projections. The pattern is what matters.
This is the cognitive-neuroscience version of norepinephrine: a precise definition, the locus coeruleus and its anatomy, the two firing modes the system uses to switch between focus and exploration, the Aston-Jones and Cohen adaptive gain theory that reframed the classic Yerkes-Dodson arousal-performance curve, what raises and lowers tonic NE in everyday life, and the drugs (caffeine, stimulants, atomoxetine, beta blockers) that work by pushing on this system. Tomatoes is a focus tool built around the kind of sustained, attentive practice the phasic mode supports. The app is free for 3 days, then $4.99/week, $29.99/year, or $39 lifetime.
What Norepinephrine Is and Where It Comes From
Norepinephrine (often abbreviated NE; the British literature uses the synonym noradrenaline and the abbreviation NA) is a catecholamine neurotransmitter, biochemically the immediate precursor of epinephrine (adrenaline) on the synthesis pathway from tyrosine. The two are close chemical cousins and historically were sometimes confused, but they do different jobs in different places. Epinephrine is mostly a hormone, released from the adrenal medulla into the bloodstream during stress and acting on heart, vasculature, and metabolism. Norepinephrine is mostly a neurotransmitter, released from neurons inside the brain and from sympathetic nerve terminals in the periphery onto specific target tissues. The same molecule can act as either, depending on where it is released.
The norepinephrine system answers the question the brain has been asking forever: how does a slow, energetically expensive cortex decide which inputs deserve processing power right now? It does this by neurochemical broadcast. A small number of neurons fire, release a modulator across vast areas, and those areas change their excitability in concert. This is fundamentally different from how, say, a visual signal travels through the brain. A visual signal moves along a specific labelled line. Norepinephrine sets the state of the whole system that the signal arrives into.
The release site that matters most for attention sits in the brainstem, in a tiny nucleus called the locus coeruleus.
The Locus Coeruleus: A Tiny Nucleus With Outsized Reach
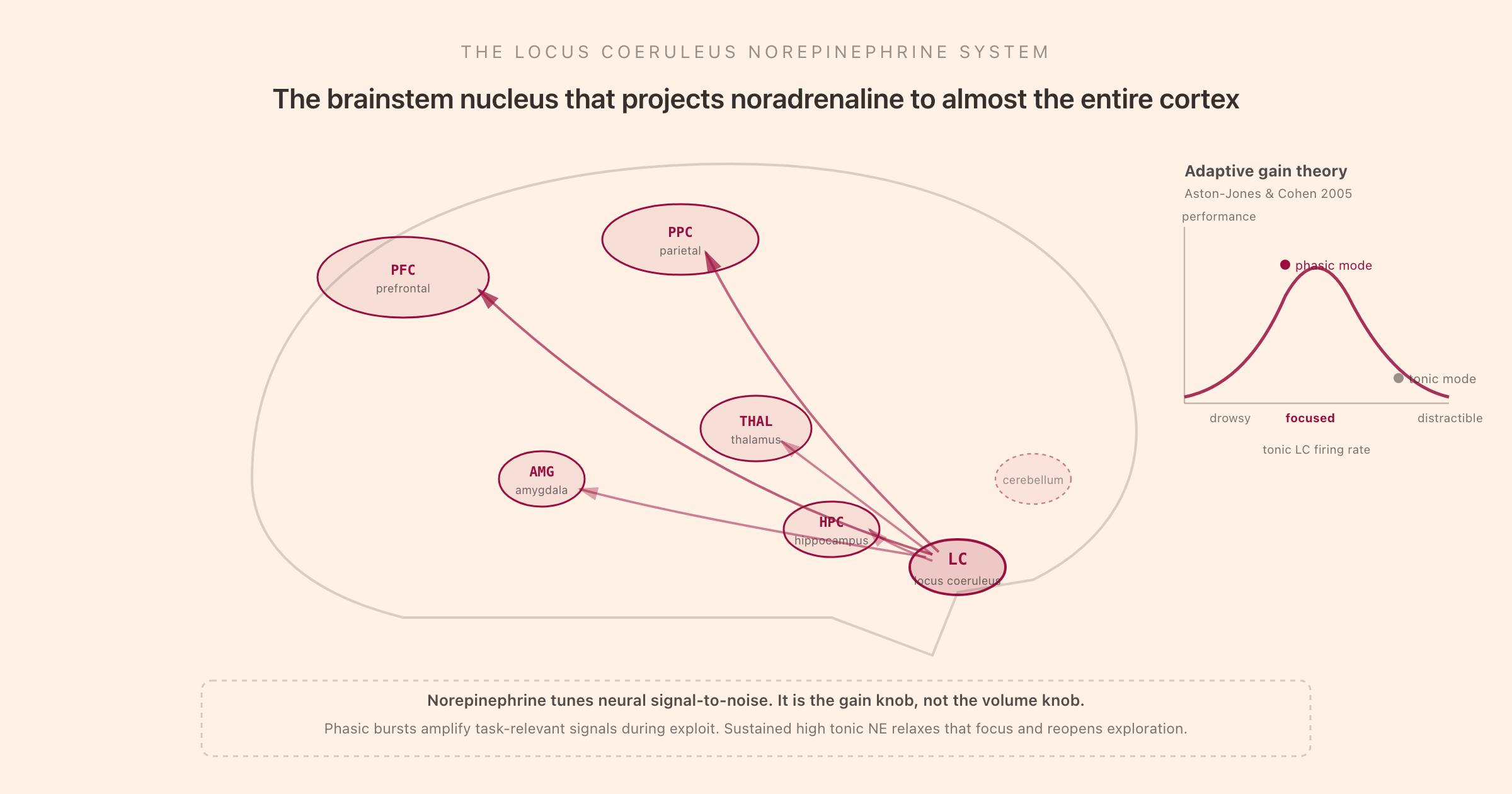
The locus coeruleus (Latin for "blue spot", named for the slight blue-green pigment its cells contain from melanin) is one of the smallest nuclei in the brain by cell count. It sits bilaterally in the dorsal pons of the brainstem, and a human LC contains on the order of 50,000 neurons. That is small, fewer than the number of neurons in a single column of cortex. What makes it consequential is what those neurons do.
Each LC neuron sends a single, profusely branching axon. The axons ascend through the medial forebrain bundle and the dorsal noradrenergic bundle, branch repeatedly, and innervate enormous territories of the brain: the entire cerebral cortex, the thalamus, the hippocampus, the amygdala, the cerebellum, and downward to the spinal cord. A single LC neuron will release norepinephrine across many millimetres of tissue. The system is built for broadcast, not point-to-point messaging. When the LC fires, large regions of the cortex receive NE almost simultaneously.
This anatomy is why norepinephrine is a global modulator rather than a circuit-specific signal. It is also why disruption of the LC affects every cognitive function the cortex supports, from working memory to perception to mood. The work of Stephen Foote, Gary Aston-Jones, Susan Sara, and colleagues over four decades established the LC's projection pattern in detail and showed that LC firing tracks behavioural state, arousal, novelty, and attentional engagement so tightly that the firing rate predicts performance moment to moment.
Phasic vs Tonic Firing: The Two Modes of the System
The single most useful idea in attention neuroscience over the past twenty years comes from the realisation that LC neurons fire in two distinct modes, and the mode matters more than the absolute rate.
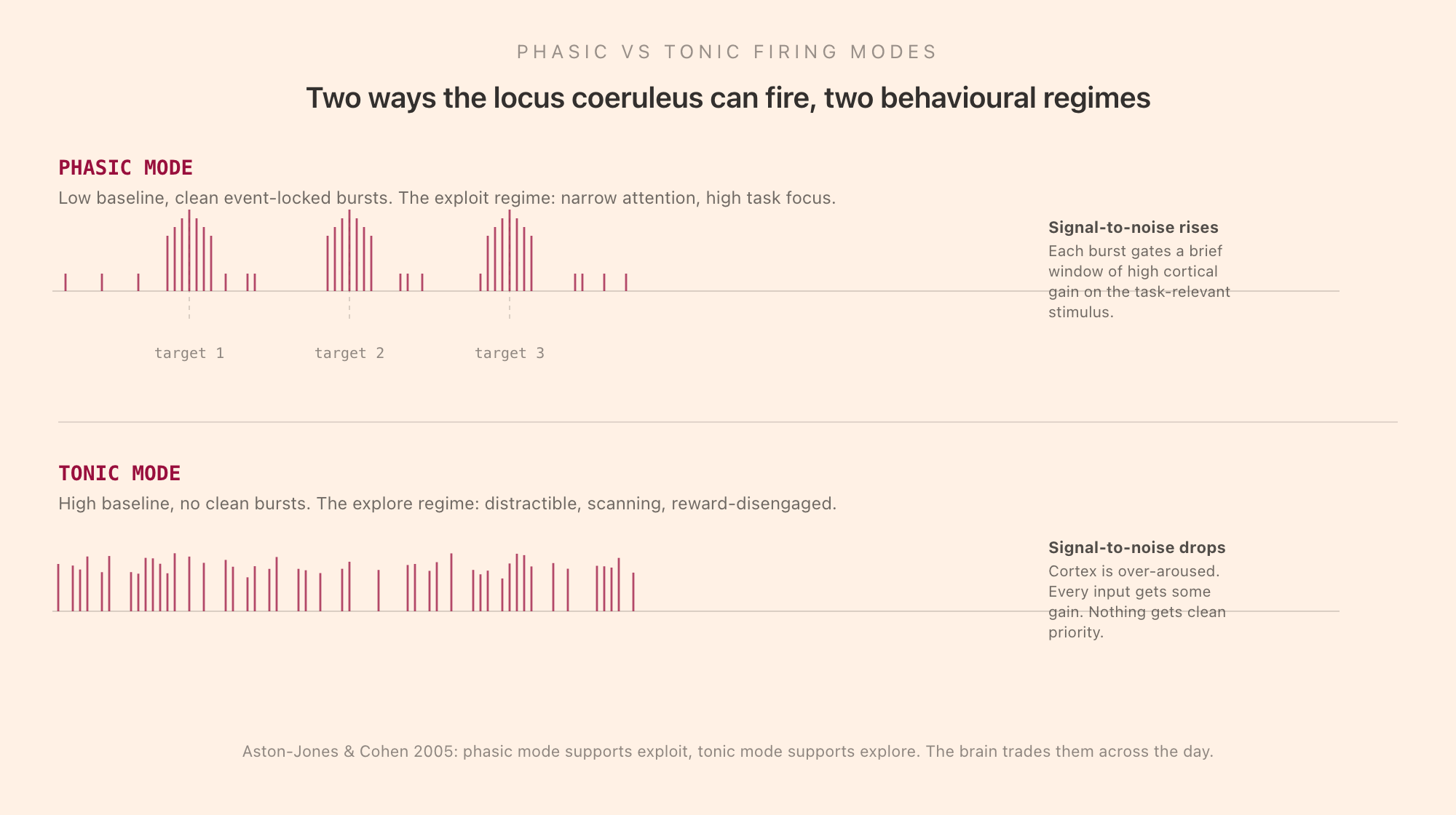
Phasic mode is what LC firing looks like during focused, on-task behaviour. The neurons sit at a low tonic baseline, between roughly 1 and 3 Hz on average, and emit sharp, brief, high-amplitude bursts of 2 to 4 spikes when a task-relevant stimulus appears. The bursts are tightly time-locked to behaviourally meaningful events: a target on a vigilance task, a reward cue, an oddball in an attended sequence. The burst lasts a few hundred milliseconds and is followed by a refractory period of relative silence. Cortically, each burst releases a pulse of NE that briefly raises the gain on cortical neurons, sharpening their response to the input that just arrived. This is what focused attention looks like at the neuromodulatory level: a clean signal carved out of low background by a precise burst.
Tonic mode is what LC firing looks like during distractible, scanning, off-task behaviour. The neurons fire at a high sustained baseline, often above 5 Hz, with no clean event-locked bursts. The bursts that arrive on a target are blunted or absent. The cortex gets a steady drizzle of NE that raises gain on everything, which sounds like it should help but does the opposite: when every input is amplified, the brain cannot prioritise. Tonic mode looks subjectively like restlessness, twitchiness, a wandering gaze that catches on anything that moves. It is the state in which an animal stops persevering on the current option and starts sampling alternatives, and Gary Aston-Jones and Jonathan Cohen proposed in 2005 that this is exactly its function.
Their adaptive gain theory is the most influential framework for understanding LC norepinephrine. The brain trades two regimes. Phasic mode supports "exploit": stick with the current task, amplify task-relevant signals, ignore distractors, finish what you started. Tonic mode supports "explore": disengage from the current task, sample the environment, find a better option. Cognitive performance on any specific task is maximised at the boundary between these regimes, with low tonic firing and strong phasic bursts. As tonic NE rises beyond that, performance falls off, the curve drops on the right side and you become distractible.
The mode is not a static trait. The same LC neurons switch between phasic and tonic regimes within seconds, dictated by reward history, novelty, and the cost-benefit calculation of staying engaged. When a task is rewarding and going well, phasic mode dominates and you focus. When rewards dry up or the task becomes painful, tonic NE rises, the cortex starts amplifying off-task inputs, and you drift. The shift is not a failure of willpower; it is the system doing what it evolved to do.
The Yerkes-Dodson Curve, Properly Understood
The classic inverted-U relationship between arousal and performance, introduced by Robert Yerkes and John Dodson in 1908 on the basis of mouse-learning experiments and now invoked in every introductory psychology textbook, is essentially a graph of tonic norepinephrine. The left descending arm is drowsiness and underarousal: tonic NE too low, cortex underexcited, performance poor. The peak is alert focused engagement: tonic NE in the right range to support clean phasic bursts. The right descending arm is over-arousal: tonic NE too high, cortex saturated, attention fragments.
What Yerkes and Dodson described phenomenologically, neuroscience eventually accounted for mechanistically. The inverted-U is the same curve adaptive gain theory predicts from tonic LC firing rate, and the empirical data from Aston-Jones, Bhavan Rajkowski, and others recording in monkeys lines up with it directly. There is a peak, the peak is narrow, and the right side falls off as fast as the left.
The textbook caveat that the optimal arousal level depends on task difficulty also turns out to track NE in a sensible way. Simple, well-learned tasks tolerate higher tonic arousal, the brain can afford to be a little keyed up and still execute them. Complex, novel tasks need lower tonic arousal and cleaner phasic bursts, because the cortical signal-to-noise advantage matters more when the work is hard. The same person who can answer emails fluently with a third coffee in them cannot do creative or technical work in that state, and the explanation is mostly that the high tonic NE is good for the easy task and bad for the hard one.
What Raises and Lowers Tonic NE in Everyday Life
The factors that move the LC across its firing modes are not mysterious; most of them are obvious once you know what you are looking at.
Stress raises tonic NE. The LC is part of the central stress response. Corticotropin-releasing factor (CRF) from the hypothalamus and the central amygdala excites LC neurons directly, pushing the system toward the high-tonic, scanning regime. Acute stress is the textbook driver of the right side of the inverted-U: rumination, twitchiness, can't sit still, can't sustain attention on one thing. The work of Rita Valentino, Elisabeth van Bockstaele, and others mapped the CRF-LC pathway in detail and showed that chronic stress can leave tonic LC firing elevated for long periods.
Novelty raises phasic NE first, then can raise tonic NE if sustained. A genuinely novel stimulus triggers a robust phasic burst in the LC, which is part of why novel things grab attention. If the environment stays unpredictable (an open-plan office, a feed of notifications, a podcast on while you work), the system shifts toward tonic mode because no single stream of inputs is winning the exploit calculation. This is the mechanism under the productivity advice to make your environment boring during deep work. A boring environment is one with no novel inputs to drive tonic NE up.
Caffeine pushes the system upward. Caffeine works primarily through adenosine receptor blockade, and one of its downstream consequences is to disinhibit LC firing. A first cup nudges tonic NE up from a low baseline (you move from the left side of the curve toward the peak, which feels good). A third cup pushes you past the peak. The dose that helps a tired person at 8 a.m. is not the dose that helps a focused person at 2 p.m.
Sleep deprivation lowers tonic NE. The LC's firing rate is tied to behavioural state; during sleep it falls dramatically, and across a sleep-deprived day it cannot maintain its waking baseline. The cognitive picture of sleep deprivation, slow, foggy, easily distracted by the wrong things, is the left-side-of-the-curve picture with episodes of compensatory over-arousal. This connects directly to the sleep stages literature: REM sleep is associated with near-total LC silence, and that quiet period appears to be necessary for sensitivity to NE to reset.
Exercise raises tonic NE acutely and improves regulation chronically. A single bout of moderate-to-vigorous aerobic exercise spikes circulating NE and central LC activity, which is part of why a workout sharpens the next hour or two of cognitive work. Chronic exercise training appears to improve the system's dynamics, the phasic bursts get cleaner against a calmer tonic baseline, which is consistent with the broader neuroplasticity literature on aerobic fitness and cognition.
Mindfulness and slow breathing lower tonic NE. Practices that downregulate sympathetic drive (slow nasal breathing, paced respiration around 5-6 breaths per minute, body scans, basic mindfulness meditation) reliably lower heart rate and circulating NE, and the central LC tracks that drop. The cognitive effect is what you'd predict: a quieter tonic baseline that lets phasic bursts do their job. Mindfulness training is not just a vibe; it is a way of moving leftward on the inverted-U when you are stuck on the right.
The Drugs That Push on This System
Several classes of widely used medication work directly through the noradrenergic system, and their clinical profiles make a lot more sense once you read them through the adaptive-gain lens.
Stimulants for ADHD (methylphenidate, amphetamines) raise synaptic NE (and dopamine) by blocking the norepinephrine transporter and, for amphetamines, by reverse-transporting NE out of vesicles into the synaptic cleft. The net effect at clinical doses is to increase phasic responsiveness and improve the signal-to-noise ratio of cortical processing. People with ADHD often sit on the left or in the over-aroused-scanning portion of the inverted-U at baseline; a well-titrated stimulant nudges them toward the peak. Too much and the right-side cliff appears: jittery, perseverative, narrowed-down focus on the wrong thing. The therapeutic window is the curve.
Atomoxetine is the cleanest example of "norepinephrine drug." It is a selective norepinephrine reuptake inhibitor with no significant dopamine action, FDA-approved for ADHD. It raises NE tonically, and its slower onset (clinical effect over weeks rather than within an hour) is consistent with adaptive changes in the LC system rather than an acute synaptic boost. It also works on cognitive control in non-ADHD populations in laboratory tasks, which is one of the cleaner pieces of evidence that NE alone, without dopamine, is enough to shift attention.
Beta blockers (propranolol) do the opposite at the periphery: they block beta-adrenergic receptors, blunting the cardiovascular effects of NE and epinephrine and breaking the somatic-feedback loop that amplifies anxiety. Centrally they have modest effects, but the well-established use case (taking a beta blocker before a performance to lower stage fright) is a clean example of cutting the right-side-of-the-curve over-arousal without sedating the brain.
Alpha-2 agonists (clonidine, guanfacine) activate inhibitory autoreceptors on LC neurons, which reduces tonic LC firing. Guanfacine is used in ADHD as an adjunct or alternative to stimulants, and its mechanism (calming the over-aroused right side of the curve, restoring cleaner phasic dynamics) is consistent with what adaptive gain theory predicts a useful pro-attention drug should do. Amy Arnsten's work in monkeys has been particularly clear about this: stress-impaired prefrontal function can be rescued by alpha-2 agonism.
SNRIs (venlafaxine, duloxetine) raise both NE and serotonin, primarily prescribed for depression and anxiety. Their cognitive profile is mixed (depression's effect on attention is multifactorial), but the NE arm of their action is implicated in the activating, attention-restoring side of antidepressant response, as distinct from the more sedating SSRI profile.
The unifying point is that norepinephrine drugs work because attention is gated by norepinephrine. The pharmacology is not a coincidence.
Norepinephrine and the Cognitive Systems Built on Top of It
The reason norepinephrine matters for focus work is that almost every cognitive system this site discusses operates at gains set by the LC. Three connections are worth making explicit.
Working memory and executive function depend on tuned NE. Amy Arnsten's lab spent decades showing that prefrontal working-memory cells need NE within a narrow window: too little and the cells cannot sustain firing across a delay; too much (the stress profile) and they go quiet. The working memory capacity ceiling is real, but how reliably you operate at that ceiling depends on whether your LC is in the right mode. The same is true of the broader executive function family of updating, inhibition, and switching.
The default mode network toggles partly through NE. The shift from on-task focused processing (task-positive networks engaged, default mode suppressed) and back to introspective wandering (default mode up, task-positive networks down) is influenced by LC firing. Phasic bursts during task engagement suppress DMN activity, supporting the default mode network on-versus-off dynamic. Sustained tonic NE without bursts is associated with the disengaged, mind-wandering profile.
Multitasking pays its switch cost partly in NE. The cost of multitasking is the cost of repeatedly resetting the cortical state. Each switch needs a fresh phasic burst to re-prime the next task's representations, and the system gets noisier when it has to do this constantly. Single-tasking is not a moral stance; it is the regime where the LC's natural exploit dynamics can actually work.
What This Tells You About Building a Focus Routine
If norepinephrine is the gain knob, then "how do I focus better" mostly means "how do I keep tonic NE in the right zone for long enough to do good work." A few practical reads fall out of the adaptive-gain picture:
- Front-load the day with the hardest work. Tonic NE is naturally higher in the morning (cortisol awakening response and circadian arousal contribute), and depletion accumulates across the day. Save the easy email and admin for the afternoon when you have less left.
- Make the environment boring for deep work. Every novel input draws a phasic burst toward off-task content, and a constant drip of novelty raises tonic NE into the distractible regime. Close tabs, hide the phone, mute the chat.
- Use Pomodoro-style timeboxing. Sustained engagement keeps the system in phasic mode, but everyone's exploit budget runs out. A short scheduled break is a controlled descent into the explore regime, after which a new pomodoro can re-establish phasic dynamics. This is one of the cleanest neuroscience reads of the pomodoro technique.
- Caffeine the right amount for the right task. A first dose pulls you toward the peak. A third dose pushes you off it. Match dose to the hardness of the work, not to how groggy you started.
- Treat sleep as the system's reset. REM sleep is when LC firing drops to near zero, which appears to restore sensitivity. A sleep-deprived day is a noradrenergically degraded day; you cannot brute-force focus through it.
- Use exercise as a pre-focus lever. A moderate aerobic block 30-60 minutes before deep work raises NE acutely and clears the day's stress baseline.
The Short Version
Norepinephrine (noradrenaline) is the brainstem-released neuromodulator that sets the gain on cortical processing. It is released from neurons in the locus coeruleus, a small bilateral nucleus in the dorsal pons that projects fibres to almost the entire cortex, the thalamus, the hippocampus, the amygdala, and the cerebellum. The system operates in two firing modes. Phasic mode (low tonic baseline plus event-locked bursts) supports focused on-task behaviour, the "exploit" regime in adaptive gain theory. Tonic mode (high sustained baseline, no bursts) supports distractible scanning, the "explore" regime. The classic Yerkes-Dodson inverted-U of arousal and performance is essentially a graph of tonic LC firing: drowsy on the left, focused at the peak, distractible on the right. Stress, caffeine, novelty, and over-arousal push tonic NE up; sleep, slow breathing, mindfulness, and rest pull it down. The drugs that improve attention (stimulants, atomoxetine, guanfacine) work through this system; the drugs that calm anxiety (beta blockers, alpha-2 agonists) work through it from the other direction. Almost every other focus-relevant system this site covers (working memory, executive function, the default mode network, multitasking) operates at gains the LC sets. Practical reads: hardest work in the morning, boring environment, timeboxed sessions, calibrated caffeine, real sleep. Tomatoes is built around the timeboxed-session piece, and is free for 3 days, then $4.99 weekly, $29.99 yearly, or $39 lifetime.